 Ce tissu sépare chacune des fibres musculaires qui composent le muscle, qui sert au soutien des élements, à l'aide au mouvment, à la protection contre les chocs et les tractions trop sévères. Ce tissu sépare chacune des fibres musculaires qui composent le muscle, qui sert au soutien des élements, à l'aide au mouvment, à la protection contre les chocs et les tractions trop sévères.
 C'est la fibre musculaire, multipliée par des milliers elle constitue le muscle. Elle est entourée d'une gaine protectrice que l'on nomme le tissu conjonctif. constituée de fibrilles, constituées de filaments faits de molécules protéiques. Leur taille à l’âge adulte est de 10 à 100 angstrom (de diamètre et jusqu’à 20cm de long. A l’origine, lors des stades de développement de l’embryon, les fibres musculaires se forment à partir de cellules encore indifférenciées nommées myoblastes qui fusionnent entre elles. Après la naissance ces fibres croissent en taille mais pas en nombre. Il y a possibilité de les regénérer par la suite après une blessure, à partir d’autres cellules indifférenciées appelées cellules satellites. Généralement, si la perte de fibres est trop importante, c’est par le biais de l’hypertrophie des fibres (augmentation de leur taille) que la compensation à lieu. Dans une myofibre, il y a plusieures myofibrilles. C'est la fibre musculaire, multipliée par des milliers elle constitue le muscle. Elle est entourée d'une gaine protectrice que l'on nomme le tissu conjonctif. constituée de fibrilles, constituées de filaments faits de molécules protéiques. Leur taille à l’âge adulte est de 10 à 100 angstrom (de diamètre et jusqu’à 20cm de long. A l’origine, lors des stades de développement de l’embryon, les fibres musculaires se forment à partir de cellules encore indifférenciées nommées myoblastes qui fusionnent entre elles. Après la naissance ces fibres croissent en taille mais pas en nombre. Il y a possibilité de les regénérer par la suite après une blessure, à partir d’autres cellules indifférenciées appelées cellules satellites. Généralement, si la perte de fibres est trop importante, c’est par le biais de l’hypertrophie des fibres (augmentation de leur taille) que la compensation à lieu. Dans une myofibre, il y a plusieures myofibrilles.
 C'est une sous-compartimentation de la cellule. Ses fonctions s'expliquent par sa capacité à délimiter un volume du reste du cytoplasme. La membrane du reticulum endoplasmique sépare la lumière du réticulum du cytosol. Dans certains types cellulaires le reticulum endoplasmique lisse spécialisé pour le stockage du calcium est appelé "réticulum sarcoplasmique" dans les cellules musculaires. Il se charge du stockage et de la concentration du calcium. C'est une sous-compartimentation de la cellule. Ses fonctions s'expliquent par sa capacité à délimiter un volume du reste du cytoplasme. La membrane du reticulum endoplasmique sépare la lumière du réticulum du cytosol. Dans certains types cellulaires le reticulum endoplasmique lisse spécialisé pour le stockage du calcium est appelé "réticulum sarcoplasmique" dans les cellules musculaires. Il se charge du stockage et de la concentration du calcium.
 Elles sont parallèles les une aux autres et allongées dans le même sens. Ce sont des molécules protéiques qui existent en deux sortes : l’actine (longue) et la myosine (courte). Une Myofibrille est constituée de sarcomères qui sont l’unité minimale de contraction musculaire. Ces sarcomères sont disposés en chaine sur toute la longueur de la fibre. Les filaments fins sont constitués de molécule actine, et les filaments épais sont faits de la molécule myosine. L’actine est une protéine aux propriétés contractiles. Elles sont parallèles les une aux autres et allongées dans le même sens. Ce sont des molécules protéiques qui existent en deux sortes : l’actine (longue) et la myosine (courte). Une Myofibrille est constituée de sarcomères qui sont l’unité minimale de contraction musculaire. Ces sarcomères sont disposés en chaine sur toute la longueur de la fibre. Les filaments fins sont constitués de molécule actine, et les filaments épais sont faits de la molécule myosine. L’actine est une protéine aux propriétés contractiles.
 Il s'agit d'un espace situé entre les Myofibrillles et le reticulum endoplasmique dans lequel se "déverse" le calcium libéré par le réticulum sarcoplasmique qui entoure les Myofibrilles. Le cytosol est limité par la membrane plasmique et l'enveloppe nucléaire et ne contient que des susbstances solubles. L'eau constitue en moyenne 85 % du cytosol. Avec ses macromolécules en suspension dans un milieux aqueux salé le cytosol présente une viscosité 4 fois supérieure à celle de l'eau et correspond à un gel colloïde. Il s'agit d'un espace situé entre les Myofibrillles et le reticulum endoplasmique dans lequel se "déverse" le calcium libéré par le réticulum sarcoplasmique qui entoure les Myofibrilles. Le cytosol est limité par la membrane plasmique et l'enveloppe nucléaire et ne contient que des susbstances solubles. L'eau constitue en moyenne 85 % du cytosol. Avec ses macromolécules en suspension dans un milieux aqueux salé le cytosol présente une viscosité 4 fois supérieure à celle de l'eau et correspond à un gel colloïde.
 Le réticulum sarcoplasmique est le nom donné au réticulum endoplasmique lisse des cellules musculaires striées squelettiques. Ce compartiment est une réserve interne d'ions Ca2+ (on l'appelle pour cette raison aussi calciosome) qui se déversent dans le cytoplasme en réponse à l'arrivée d'une dépolarisation de la membrane plasmique causée par la fixation de l'ACh sur des récepteurs nicotiniques de la cellule musculaire. Cette augmentation de calcium cytoplasmique provoque la contraction musculaire en permettant l'interaction entre la myosine et les microfilaments d'actine. Le calcium est ensuite repompé (grâce à une pompe consommant de l'ADP) dans le réticulum sarcoplasmique. Le réticulum sarcoplasmique déploie sont réseau de manière particulière entre la membrane cytoplasmique et le sarcomère de telle sorte qu'un potentiel d'action se propage le long du réticulum sarcoplasmique, et que le calcium libéré soit directement en contact avec le sarcomère pour initier la contraction. Le réticulum sarcoplasmique est le nom donné au réticulum endoplasmique lisse des cellules musculaires striées squelettiques. Ce compartiment est une réserve interne d'ions Ca2+ (on l'appelle pour cette raison aussi calciosome) qui se déversent dans le cytoplasme en réponse à l'arrivée d'une dépolarisation de la membrane plasmique causée par la fixation de l'ACh sur des récepteurs nicotiniques de la cellule musculaire. Cette augmentation de calcium cytoplasmique provoque la contraction musculaire en permettant l'interaction entre la myosine et les microfilaments d'actine. Le calcium est ensuite repompé (grâce à une pompe consommant de l'ADP) dans le réticulum sarcoplasmique. Le réticulum sarcoplasmique déploie sont réseau de manière particulière entre la membrane cytoplasmique et le sarcomère de telle sorte qu'un potentiel d'action se propage le long du réticulum sarcoplasmique, et que le calcium libéré soit directement en contact avec le sarcomère pour initier la contraction.
 Organites responsables de la respirations: remplies d'enzimes qui transforment les sucres et les graisses en ATP avec l'aide d'oxygène. Elles oxydent donc le carbone pour fournir de l’energie. C’est à ce niveau qu’ont lieu les réactions d’oxydation du carbonne qui produisent de l’ATP. C’est là qu’a lieu l’oxydation de la molécule de glucose. En fait le glucose est oxydé dans le cytoplasme : il devient de petites molécules carbonnée, nommées « pyruvate ». le pyruvate est transporté dans la matrice mitochondriale, où il est privé progressivement de ses composés carbone et hydrogène pour créer, entre autes, de l’ATP et du CO2. Organites responsables de la respirations: remplies d'enzimes qui transforment les sucres et les graisses en ATP avec l'aide d'oxygène. Elles oxydent donc le carbone pour fournir de l’energie. C’est à ce niveau qu’ont lieu les réactions d’oxydation du carbonne qui produisent de l’ATP. C’est là qu’a lieu l’oxydation de la molécule de glucose. En fait le glucose est oxydé dans le cytoplasme : il devient de petites molécules carbonnée, nommées « pyruvate ». le pyruvate est transporté dans la matrice mitochondriale, où il est privé progressivement de ses composés carbone et hydrogène pour créer, entre autes, de l’ATP et du CO2.
 Unité minimale de contraction musculaire. C’est le sous-ensemble : les sarcomères se rétractent chacun leur tour. Il n’y a aucun changement de longueur de sarcomère lors d’une contraction (et donc aucun changement de longueur du muscle) : ce sont des coulissements. Lors d’une contraction, il y a réduction d’environ 20% de la longueur du sarcomère par effet de coulissage des fibrilles. La vitesse de contraction est de 15 angstrom/seconde. Plus un muscle est divisé en sarcomères plus il donnera l’impression de retrécir. Les filaments fins (actine) coulissent le long des filaments plus épais (myosine). Lors du raccourcissement du sarcomère, il n’y a pas de raccourcissement des filaments fins ou épais, mais un glissement qui fait qu’ils se chevauchent. C’est l’image de l’aviron. la myosine est dotée de ponts traversaux qui viennent s’attacher à la molécule d’actine. Au centre du sarcomère se situe la molécule de myosine, entourée de part et d’autre de six molécules d’actine. La molécule de myosine va coulisser en s’appuyant sur les molécules d’actine. A l’extrémité du sarcomère (de chaque coté) se situe une « ligne Z » qui permet aux molécules d’actine de se connecter, liant ainsi chaque sarcomère avec le suivant et le précédant. Le chevauchement est un peu plus compliqué en fait : chaque filament épais est entouré de six filaments fins disposés en hexagone. Chaque filament fin est donc entouré à son tour de trois filaments épais en triangles. C’est au niveau de l’espace entre ces filaments qu’à lieu le phénomène de contraction musculaire au niveau des « ponts traversaux». Les « ponts traversaux » des molécules de myosine viennent s’arrimer aux têtes des molécules d’actine pour y coulisser. C’est en s’appuyant sur les filaments fins que les filaments épais coulissent. Une Myofibrille est composée de deux grands types de molécules : la myosine et l’actine. Unité minimale de contraction musculaire. C’est le sous-ensemble : les sarcomères se rétractent chacun leur tour. Il n’y a aucun changement de longueur de sarcomère lors d’une contraction (et donc aucun changement de longueur du muscle) : ce sont des coulissements. Lors d’une contraction, il y a réduction d’environ 20% de la longueur du sarcomère par effet de coulissage des fibrilles. La vitesse de contraction est de 15 angstrom/seconde. Plus un muscle est divisé en sarcomères plus il donnera l’impression de retrécir. Les filaments fins (actine) coulissent le long des filaments plus épais (myosine). Lors du raccourcissement du sarcomère, il n’y a pas de raccourcissement des filaments fins ou épais, mais un glissement qui fait qu’ils se chevauchent. C’est l’image de l’aviron. la myosine est dotée de ponts traversaux qui viennent s’attacher à la molécule d’actine. Au centre du sarcomère se situe la molécule de myosine, entourée de part et d’autre de six molécules d’actine. La molécule de myosine va coulisser en s’appuyant sur les molécules d’actine. A l’extrémité du sarcomère (de chaque coté) se situe une « ligne Z » qui permet aux molécules d’actine de se connecter, liant ainsi chaque sarcomère avec le suivant et le précédant. Le chevauchement est un peu plus compliqué en fait : chaque filament épais est entouré de six filaments fins disposés en hexagone. Chaque filament fin est donc entouré à son tour de trois filaments épais en triangles. C’est au niveau de l’espace entre ces filaments qu’à lieu le phénomène de contraction musculaire au niveau des « ponts traversaux». Les « ponts traversaux » des molécules de myosine viennent s’arrimer aux têtes des molécules d’actine pour y coulisser. C’est en s’appuyant sur les filaments fins que les filaments épais coulissent. Une Myofibrille est composée de deux grands types de molécules : la myosine et l’actine.
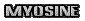 La déformation de la tête de myosine a une amplitude correspondant au diamètre d’une molécule d’actine. Ce basculement est rendu possible par l’hydrolyse d’une molécule d’ATP qui vient se fixer sur la tête de myosine. Chaque molécule de myosine est dotée d’une « queue » qui court le long de l’axe dotée aux extrémités de deux « têtes » globulaires (ce sont les « ponts traversaux). Il y a donc deux globules pour la fixation : un pour la fixation de l’ATP et l’autre pour la fixation de l’actine. Le pont traversal est situé de chaque coté de l’extrémité de la molécule de myosine et « tire » la molécule d’actine vers l’intérieur, raccoucissant ainsi le sarcomère. La déformation de la tête de myosine a une amplitude correspondant au diamètre d’une molécule d’actine. Ce basculement est rendu possible par l’hydrolyse d’une molécule d’ATP qui vient se fixer sur la tête de myosine. Chaque molécule de myosine est dotée d’une « queue » qui court le long de l’axe dotée aux extrémités de deux « têtes » globulaires (ce sont les « ponts traversaux). Il y a donc deux globules pour la fixation : un pour la fixation de l’ATP et l’autre pour la fixation de l’actine. Le pont traversal est situé de chaque coté de l’extrémité de la molécule de myosine et « tire » la molécule d’actine vers l’intérieur, raccoucissant ainsi le sarcomère.
 Molécule filamentaire dont le déplacement vers le long de la molécule de myosine vers son milieu constitue l'élément clé du raccourcissement du sarcomère, initiant ainsi la contraction musculaire. Une molécule d'actine acceuille la molécule de tropomyosine qui s'enroule autour d'elle (bloquant les têtes d'actine), et les molécules de troponine (de forme globulaire) qui lient l'actine et la tropomyosine et peuvent déplacer cette dernière quand elles changent de position après avoir accueilli un ion calcium. Après que l’ion calcium ait modifié son extrémité, elle reçoit la tête de myosine de la fibrile voisine qui se loge dans la cavité de la tête d’actine. la molécule d’actine est une protéine globulaire qui s’associe avec d’autres molécules d’actine pour former des chaines. Cette chaine a une forme hélicoidale, et constitue un « filament fin ». Chaque molécule d’actine possède une « tête » de fixation pour la myosine. Une filament fin possède donc plusieures tetes de fixation. Molécule filamentaire dont le déplacement vers le long de la molécule de myosine vers son milieu constitue l'élément clé du raccourcissement du sarcomère, initiant ainsi la contraction musculaire. Une molécule d'actine acceuille la molécule de tropomyosine qui s'enroule autour d'elle (bloquant les têtes d'actine), et les molécules de troponine (de forme globulaire) qui lient l'actine et la tropomyosine et peuvent déplacer cette dernière quand elles changent de position après avoir accueilli un ion calcium. Après que l’ion calcium ait modifié son extrémité, elle reçoit la tête de myosine de la fibrile voisine qui se loge dans la cavité de la tête d’actine. la molécule d’actine est une protéine globulaire qui s’associe avec d’autres molécules d’actine pour former des chaines. Cette chaine a une forme hélicoidale, et constitue un « filament fin ». Chaque molécule d’actine possède une « tête » de fixation pour la myosine. Une filament fin possède donc plusieures tetes de fixation.
Enroulés autour de la molécule d’actine il y a les molécules de troponine et de tropomyosine qui rendent possible ou bloquent la cohésion actine-myosine.
La tropomyosine est une molécule constituée de deux fins filaments enroulée autour de la molécule d’actine.
La troponine, de forme globulaire, est à la fois fixée à la tropomyosine et à l’actine.
La troponine fonctionne comme un roulement qui fait se déplacer le « cable » de tropomyosine sur l’actine.
La tropomyosine recouvre une partie des têtes de fixation de l’actine, empêchant ainsi les ponts traversaux de la myosine de se fixer à l’actine, et donc la contraction musculaire d’avoir lieu.
Pour que la contraction commence il faut que la troponine déplace-débloque la tropomyosine en la faisant bouger latéralement pour qu’elle libère les sites d’ancrage qui accueillent la myosine. Pour que la troponine se déplace il faut que du calcium vienne s’y fixer. Quand le calcium y est fixé, la forme générale de la molécule de troponine est modifiée et cette dernière se déplace, entraînant ainsi le filament de tropomyosine, qui libère à son tour le têtes de fixation de l’actine, permettant ainsi à la myosine de venir s’y fixer. Quand le calcium quitte la troponine cette dernière reprend sa place naturelle, et « tire » avec elle la tropomyosine qui vient se placer devant les sites de fixation de la myosine, et empêche de nouveau la contraction d’avoir lieu.
 C'est la molécule qui, en se déplaçant, libère ou occulte les têtes d'actine en entraînant dans son mouvement la molécule filamentaire de tropomyosine. Le fonctionnement est proche de celui de la serrure. Quand une molécule de calcium vient s'arrimer à la molécule de troponine elle colisse (comme un barillet de serrure) et ce mouvement déplace la molécule de tropomyosine (tel un loquet qui bloquait une porte) et libère le site de fixation des ponts traversaux de la molécule de myosine. C'est la molécule qui, en se déplaçant, libère ou occulte les têtes d'actine en entraînant dans son mouvement la molécule filamentaire de tropomyosine. Le fonctionnement est proche de celui de la serrure. Quand une molécule de calcium vient s'arrimer à la molécule de troponine elle colisse (comme un barillet de serrure) et ce mouvement déplace la molécule de tropomyosine (tel un loquet qui bloquait une porte) et libère le site de fixation des ponts traversaux de la molécule de myosine.
 Il s'agit d'une longue molécule qui a pour fonction de servir de vérou à la contraction . Cette molécule enroulée autour du filament fin est déplacée par les mouvements de la molécule de troponine et se positionne soit desssus, soit à coté, de la tête d'actine qui a pour fonction de permettre à la myosine de venir s'y fixer. La tropomyosine est donc l'équivalent biologique du cache-prise ou du loquet, ou plus simplement, du bout de plastique que l'on poserait sur un aimant pour empêcher la connection. Il s'agit d'une longue molécule qui a pour fonction de servir de vérou à la contraction . Cette molécule enroulée autour du filament fin est déplacée par les mouvements de la molécule de troponine et se positionne soit desssus, soit à coté, de la tête d'actine qui a pour fonction de permettre à la myosine de venir s'y fixer. La tropomyosine est donc l'équivalent biologique du cache-prise ou du loquet, ou plus simplement, du bout de plastique que l'on poserait sur un aimant pour empêcher la connection.
 Il s'agit de la partie de la molécule myosine qui effectue le travail d'ancrage-tirage-décrochage avec la tête de la molécule d'actine pour créer le racourcissement du sarcomère et ainsi générer une force. Ce pont traversal a besoin de l'ATP qu'il dégrade pour pouvoir se déplacer et aller chercher la tête de myosine, puis d'ATP encore pour se décrocher. Cycle des ponts traversaux : unité de mouvement musculaire : fixation du pont traversal sur la tête d’actine, déplacement, détachement du pont traversal, déplacement du pont pour attraper une autre tête d’actine. Il s'agit de la partie de la molécule myosine qui effectue le travail d'ancrage-tirage-décrochage avec la tête de la molécule d'actine pour créer le racourcissement du sarcomère et ainsi générer une force. Ce pont traversal a besoin de l'ATP qu'il dégrade pour pouvoir se déplacer et aller chercher la tête de myosine, puis d'ATP encore pour se décrocher. Cycle des ponts traversaux : unité de mouvement musculaire : fixation du pont traversal sur la tête d’actine, déplacement, détachement du pont traversal, déplacement du pont pour attraper une autre tête d’actine.
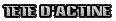 C'est le site où a lieu la connexion entre les ponts traversaux de la myosine et l'actine. C'est à ce niveau que s'accroche la myosine pour ensuite "tirer" vers l'intérieur les molécules d'actine, raccourcissant ainsi le sarcomère et créant une force de traction. Ces têtes d'actine sont généralement occultées par une molécule nommée tropomyosine, ce qui empêche la mise en oeuvre de la contraction. C'est le site où a lieu la connexion entre les ponts traversaux de la myosine et l'actine. C'est à ce niveau que s'accroche la myosine pour ensuite "tirer" vers l'intérieur les molécules d'actine, raccourcissant ainsi le sarcomère et créant une force de traction. Ces têtes d'actine sont généralement occultées par une molécule nommée tropomyosine, ce qui empêche la mise en oeuvre de la contraction.
 C'est le nom donné à la division de la myofibre correspondant au sarcomère. C'est le nom donné à la division de la myofibre correspondant au sarcomère.
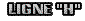 Il s'agit de la zone centrale du sarcomère : c'est un espace où il n’y a que de la myosinevers laquelle se déplace l’actine. Chaque molécule de myosine est entourée de molécules d'actine à ses deux extrémités, quand la myosine fait coulisser les molécules d'actine ce mouvement a lieu en direction du centre de la molécule de myosine, dans la zone nommée "Ligne H". Il s'agit de la zone centrale du sarcomère : c'est un espace où il n’y a que de la myosinevers laquelle se déplace l’actine. Chaque molécule de myosine est entourée de molécules d'actine à ses deux extrémités, quand la myosine fait coulisser les molécules d'actine ce mouvement a lieu en direction du centre de la molécule de myosine, dans la zone nommée "Ligne H".
 Il s'agit de la zone contraire à la ligne H, c'est à dire celle ou il n'y a que des molécles d'actine; c'est à ce niveau qu'ont lieu les déplacements. Il s'agit de la zone contraire à la ligne H, c'est à dire celle ou il n'y a que des molécles d'actine; c'est à ce niveau qu'ont lieu les déplacements.
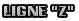 C'est la zone située entre deux sarcomères : c’est là où a lieu l’elasticité du muscle. Quand un muscle s'étire sous le poids d'une charge qui dépasse la capacité contractile du muscle, ce ne sont pas les sarcomères qui s'étirent mais la zone située entre eux qui a une capacité élastique; la "Zone Z". C'est la zone située entre deux sarcomères : c’est là où a lieu l’elasticité du muscle. Quand un muscle s'étire sous le poids d'une charge qui dépasse la capacité contractile du muscle, ce ne sont pas les sarcomères qui s'étirent mais la zone située entre eux qui a une capacité élastique; la "Zone Z".
 Le calcium est un ion fondamental pour le fonctionnement cellulaire. Il permet donc d’ « ouvrir » la porte de fixation de la molécule d’actine pour acceuillir la tête de la molécule de myosine, ouverture qui a lieu quand le calcium modifie la forme de la petite molécule de troponine et la fait ainsi se déplacer, ce qui engendre le déplacement de la molécule de tropomyosine qui bloquait l’accès à la tête de fixation de l’actine. Le calcium est un ion fondamental pour le fonctionnement cellulaire. Il permet donc d’ « ouvrir » la porte de fixation de la molécule d’actine pour acceuillir la tête de la molécule de myosine, ouverture qui a lieu quand le calcium modifie la forme de la petite molécule de troponine et la fait ainsi se déplacer, ce qui engendre le déplacement de la molécule de tropomyosine qui bloquait l’accès à la tête de fixation de l’actine.
La calcium vient se loger dans le cytoplasme pour activer le mouvement des ponts traversaux. Tout ce joue au niveau de l’actine, plus particulièrement au niveau des deux protéines qui constituent le filament d’actine : la troponine (boule) et la tropomyosine (filament). Ce sont des phénomènes electriques qui sont à l’origine de la fixation du calcium sur la troponine, et de sa libération.
Le calcium provient d’une « gaine » (ou manchons) entourant les Myofibrilles (le réticulum sarcoplasmique). A l’intérieur de ces manchons se trouvent des zones de stockage de calcium. Le calcium se trouve soit dans le réticulum sarcoplasmique, soit dans le cytosol.
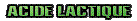 L'acide lactique est le sous-produit de la consommation par les muscles du glycogène. La glycolyse (consommation de sucre par le muscle pour faire de l'énergie) peut se faire avec un apport suffisant d'oxigène (aérobie) ou insuffisant (anaérobie). En aérobie cette consommation a lieu des les mitochondries qui consommeront totalement l'acide lactique indirectement créé. Dans l'effort intense où l'apport en oxygène est insuffisant la consommation du sucre a lieu dans le cytoplasme, et l'acide lactique ne peut être éliminé par la respiration. L'acide lactique est le sous-produit de la consommation par les muscles du glycogène. La glycolyse (consommation de sucre par le muscle pour faire de l'énergie) peut se faire avec un apport suffisant d'oxigène (aérobie) ou insuffisant (anaérobie). En aérobie cette consommation a lieu des les mitochondries qui consommeront totalement l'acide lactique indirectement créé. Dans l'effort intense où l'apport en oxygène est insuffisant la consommation du sucre a lieu dans le cytoplasme, et l'acide lactique ne peut être éliminé par la respiration.
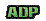 Quand une molécule d'ATP est hydrolysée (dissoute par l'eau) elle se "casse" en un élément ADP et un élément phosphate, et libère de l'énergie. Le recouplage de ces deux parties permet de reformer de l'ATP, mais cela va consommer la même dose d'énergie. Quand une molécule d'ATP est hydrolysée (dissoute par l'eau) elle se "casse" en un élément ADP et un élément phosphate, et libère de l'énergie. Le recouplage de ces deux parties permet de reformer de l'ATP, mais cela va consommer la même dose d'énergie.
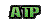 Molécule dont les liaisons entre les atomes sont très énergétiques. C'est un composé phosphaté riche en énergie. C'est la molécule qui sert à produire de l'énergie. Sa dégradation par l'eau (hydrolyse) libère de l'énergie (environ 30kj/mol), et sa synthèse consomme la même quantité d'énergie. L'hydrolyse de l'ATP donne une molécule d'ADP et un composé phosphate.. Molécule dont les liaisons entre les atomes sont très énergétiques. C'est un composé phosphaté riche en énergie. C'est la molécule qui sert à produire de l'énergie. Sa dégradation par l'eau (hydrolyse) libère de l'énergie (environ 30kj/mol), et sa synthèse consomme la même quantité d'énergie. L'hydrolyse de l'ATP donne une molécule d'ADP et un composé phosphate..
 Neurotranmeteur. Neurotranmeteur.
 Action de l'eau sur la molécule qui coupe la molécule et qui libère de l'energie chimique dans le cas de l'ATP. On casse pour créer de l'énergie. Action de l'eau sur la molécule qui coupe la molécule et qui libère de l'energie chimique dans le cas de l'ATP. On casse pour créer de l'énergie.
 Quand un composé comme l'ADP devient de l'ATP. C'est le contraire de l'hydrolise. On utilise de l'énergie pour réassembler. Quand un composé comme l'ADP devient de l'ATP. C'est le contraire de l'hydrolise. On utilise de l'énergie pour réassembler.
 Perte d'électron de l'ATP captée par une autre molécule. Perte d'électron de l'ATP captée par une autre molécule.
 Perte d'électrons (devient plus), gain d'électrons (devient moins). Perte d'électrons (devient plus), gain d'électrons (devient moins).
 Ce sont elles qui sont responsables du « message » de libération du Calcium. Les motoneurones sont des cellules nerveuses dont les axones se divisent en branches au contact du muscle et se connectent à un maximum de Myofibrilles. 1 neuronne ennerve plusieures fibres. L’ensemble Motoneurone+myofibre se nomme « unité motrice ». Il est interessant de remarquer que les myofibres connectées à un motoneurone ne sont pas forcément cote-à-cote. Plus il y a d’axones, plus il y aura de myofibres qui recevront le message de libération de calcium, et plus il y aura de contractions de sarcomères, et donc plus puissante sera la contraction. C’est pourquoi certaines personnes d’apparence mince sont très puissante, car ils sont capables de mobiliser plus de fibrilles que d’autres. Rien ne sert d’avoir des muscles hypertrophiés si nos connexions nerveuses ne sont pas capables de les stimuler. Au niveau de la myofibre, dans les sillons à la surface de la membrane plasmique (qui protègle les Myofibrilles) se situe une « plaque motrice » où vient se loger l’axone moteur (la terminaison du neurone). Cette jonction est nommée « Jonction neuromusculaire). Quand le « potenteil d’action » entre en contact avec la membrane plasmique, et se propage le long de la surface de la fibre musculaire. Au niveau des terminaisons des axones se trouvent des vésicules qui libèrent un neurotransmetteur (Ach). Quand un potentiel d’action entre en contact avec la plaque motrice il dépolarise la membrane plasmique ouvrant ainsi des canaux qui vont libérer le calcium. Ces canaux sont dits « potentiel-dépendants » car leur ouverture est liée à l’impulsion d’un potentiel d’action. Ce calcium libéré permet la libération du neurotransmetteur. Ce sont elles qui sont responsables du « message » de libération du Calcium. Les motoneurones sont des cellules nerveuses dont les axones se divisent en branches au contact du muscle et se connectent à un maximum de Myofibrilles. 1 neuronne ennerve plusieures fibres. L’ensemble Motoneurone+myofibre se nomme « unité motrice ». Il est interessant de remarquer que les myofibres connectées à un motoneurone ne sont pas forcément cote-à-cote. Plus il y a d’axones, plus il y aura de myofibres qui recevront le message de libération de calcium, et plus il y aura de contractions de sarcomères, et donc plus puissante sera la contraction. C’est pourquoi certaines personnes d’apparence mince sont très puissante, car ils sont capables de mobiliser plus de fibrilles que d’autres. Rien ne sert d’avoir des muscles hypertrophiés si nos connexions nerveuses ne sont pas capables de les stimuler. Au niveau de la myofibre, dans les sillons à la surface de la membrane plasmique (qui protègle les Myofibrilles) se situe une « plaque motrice » où vient se loger l’axone moteur (la terminaison du neurone). Cette jonction est nommée « Jonction neuromusculaire). Quand le « potenteil d’action » entre en contact avec la membrane plasmique, et se propage le long de la surface de la fibre musculaire. Au niveau des terminaisons des axones se trouvent des vésicules qui libèrent un neurotransmetteur (Ach). Quand un potentiel d’action entre en contact avec la plaque motrice il dépolarise la membrane plasmique ouvrant ainsi des canaux qui vont libérer le calcium. Ces canaux sont dits « potentiel-dépendants » car leur ouverture est liée à l’impulsion d’un potentiel d’action. Ce calcium libéré permet la libération du neurotransmetteur.
L’Ach se fixe sur la plaque motrice et transmet le potentiel d’action. La plaque motrice porte une enzyme (la choline) qui va profiter de la connexion pour remonter vers la terminaison axonique et être utilisée pour créer une nouvelle Ach. L’activité electrique de la membrane plasmique agit indirectement sur les protéines contractiles en libérant du calcium qui se répand dans le cytosol. Ce calcium est stocké dans le réticulum sarcoplasmique au niveau des « citernes terminales ».
 C'est la partie terminale nue (non protégée par une gaine) du neurone moteur, celle qui transmet les informations par le biais d'un neurotransmetteur. A l'instar d'un cable electrique, il s'agirait de la partie terminale où le fil est dénudé. C'est la partie terminale nue (non protégée par une gaine) du neurone moteur, celle qui transmet les informations par le biais d'un neurotransmetteur. A l'instar d'un cable electrique, il s'agirait de la partie terminale où le fil est dénudé.
 C'est la zone sur laquelle se fait la connection entre le système nerveux et le système musculaire. Cette zone est située au niveau de la myofibre, dans les sillons de sa surface irrégulière (membrane plasmique). C'est à cet endroit que se place la terminaison du motoneurone (l'axone moteur). C'est à ce niveau qu'ont lieu les échanges de neurotransmetteurs qui créent le potentiel d'action menant à la libération du calciumà l'intérieur du cytosol et au début de la contraction. A l'instar d'une connection électrique domestique, il s'agirait du "contact". C'est la zone sur laquelle se fait la connection entre le système nerveux et le système musculaire. Cette zone est située au niveau de la myofibre, dans les sillons de sa surface irrégulière (membrane plasmique). C'est à cet endroit que se place la terminaison du motoneurone (l'axone moteur). C'est à ce niveau qu'ont lieu les échanges de neurotransmetteurs qui créent le potentiel d'action menant à la libération du calciumà l'intérieur du cytosol et au début de la contraction. A l'instar d'une connection électrique domestique, il s'agirait du "contact".
 C'est le nom donné à l'ensemble Motoneurone-myofibre: puisque chaque neurone moteur est connecté à une fibre musculaire, la contraction globale du muscle est le fruit de l'action simultanée de ces réseaux indépendants: plusieurs motoneurones vont stimuler simultanément la myofibreà laquelle ils sont rattachés et la somme des stimulations-contractions de chacune de ces unités motrices donne la force. Comparé à un moteur à explosion, l'unité motrice correspondrait dans l'esprit à l'assemblage bougie-chambre de combustion; la bougie est le motoneurone apportant l'information, et la myofibre la chambre de combustion où ont lieu les réactions chimiques menant au développement d'une force. Le travail simultané de chaque système bougie-chambre (unité motrice) donne le mouvement. C'est le nom donné à l'ensemble Motoneurone-myofibre: puisque chaque neurone moteur est connecté à une fibre musculaire, la contraction globale du muscle est le fruit de l'action simultanée de ces réseaux indépendants: plusieurs motoneurones vont stimuler simultanément la myofibreà laquelle ils sont rattachés et la somme des stimulations-contractions de chacune de ces unités motrices donne la force. Comparé à un moteur à explosion, l'unité motrice correspondrait dans l'esprit à l'assemblage bougie-chambre de combustion; la bougie est le motoneurone apportant l'information, et la myofibre la chambre de combustion où ont lieu les réactions chimiques menant au développement d'une force. Le travail simultané de chaque système bougie-chambre (unité motrice) donne le mouvement.
 Ce que l'on nomme généralement "l'influx nerveux" est en fait constitué de potentiels d'action. Cela correspond à un changement de polarité de la cellule. Ce sont les ions (potassium, sodium) qui sont à l'origine de la différence de potentiel électrique entre l'intérieur et l'extérieur de la cellule. Le potentiel d'action est constitué d'une succession d'événements : une dépolarisation transitoire et locale de cet état de repos, d'une amplitude (agmentation du potentiel de la membrane internet). une repolarisation de la membrane interne. Le potentiel d'action dure entre 2 et 3 millisecondes. Ce que l'on nomme généralement "l'influx nerveux" est en fait constitué de potentiels d'action. Cela correspond à un changement de polarité de la cellule. Ce sont les ions (potassium, sodium) qui sont à l'origine de la différence de potentiel électrique entre l'intérieur et l'extérieur de la cellule. Le potentiel d'action est constitué d'une succession d'événements : une dépolarisation transitoire et locale de cet état de repos, d'une amplitude (agmentation du potentiel de la membrane internet). une repolarisation de la membrane interne. Le potentiel d'action dure entre 2 et 3 millisecondes.
|



















